

DID YOU KNOW? Vitamin A and beta-carotene are vital for reproductive processes like ovulation.
20 Jul 2026
Account Manager, Michelle Lutz, provides an overview of the benefits of vitamin A and beta-carotene for reproductive performance in cattle.

Amidst the delayed stabilization of the US cattle herd inventory, feeder cattle value escalated to an all-time high at around $328/cwt (CME Feeder Cattle Index, 2025), rendering producers more vulnerable to the impact of morbidity and production losses. Feedlot diets today differ in complexity to those fed in the early 1960’s (Engle et al., 2025) and include multiple milling by-products as opposed to a simple corn silage and grain diet. In essence, we are feeding a very different animal today with different breed influences, longer days on feed, and to a much heavier end weight with carcasses nearing 1000 lbs. Coupled with the genetic, nutritional, feed bunk management and feed delivery advances made in our industry over the last few decades, we now have a more efficient and productive animal with increased nutrient requirements.
Vitamin nutrition, albeit an essential nutrient (NRC, 1996) and updated guidelines for supplementation with a clear link to how it supports health, and productivity has lagged in terms of research and its consistent inclusion in diets for high performing animals. Comparing industry practice by way of the Feedlot Consulting Nutritionist Survey (Samuelson et al.,2016) to NASEM recommendations for finishing cattle (NASEM, 2000), industry practice reflects supplementing Vitamin A at levels similar to NASEM recommendations at 2,000 IU/kg DM vs 2,200 IU/kg DM; Vitamin D at only 142 IU/kg DM (mode at 0 IU/kg DM) compared to NASEM’s 275 IU/kg DM and Vitamin E at an average of 25 IU (mode of 0 IU/kg DFM) compared to NASEM’s 50-100 IU/kg DM. The most recent publication of the optimum vitamin nutrition guidelines (OVN, dsm-firmenich, 2022) for vitamins A, D and E exceed that of NASEM and industry practice for nearly all classes of beef cattle, arguing optimized health and productivity in lieu of merely avoiding deficiency.
Vitamins are essential for their role in controlling animal metabolism and facilitating nutrient synthesis and degradation and should be considered when making any dietary adjustments due to their wide range of interactions with other nutrients. Stemming from the aforementioned factors and a lack of new nutritional advancements and existing feed additives failing to deliver consistent, measurable improvements in health and weight gain, nutritionists are focusing on the fundamentals of nutrition, critically assessing diets for all nutrients but vitamins A, D and E specifically to mitigate morbidity and support weight gain.
Vitamin Requirements
Beef cattle with a fully functional rumen mainly require supplementation of the fat-soluble vitamins A, D and E (Figure 1) which are not synthesized in the rumen (NRC, 2000). Significant quantities of vitamin K are synthesized by the ruminal bacterial flora and abundant in pasture and green forages rendering supplementation unnecessary. Due to extensive ruminal synthesis of all water-soluble B-vitamins such as vitamin B12, thiamin, niacin and choline, supplementation is deemed unnecessary (NASEM, 2000, NRC, 1996, Engle, 2025). Exceptions would be thiamin supplementation at 120 to 240 mg/kg reducing the incidence of Polyencephalomalacia, induced by either a thiamin deficiency or hydrogen sulfide gas formation due to high dietary (e.g. distillers-based diets) or water sulfur levels (Ward and Patterson, 2004; Olkowski et al., 1992; Neville et al., 2010; Engle et al., 2025). Ruminal synthesis of vitamin B12 (cobalamin) is sufficient under normal circumstances but due to its role in propionate metabolism could become limiting with insufficient dietary intake of cobalt as 3 to 13% of Co is converted to vitamin B12 (Smith, 1997).
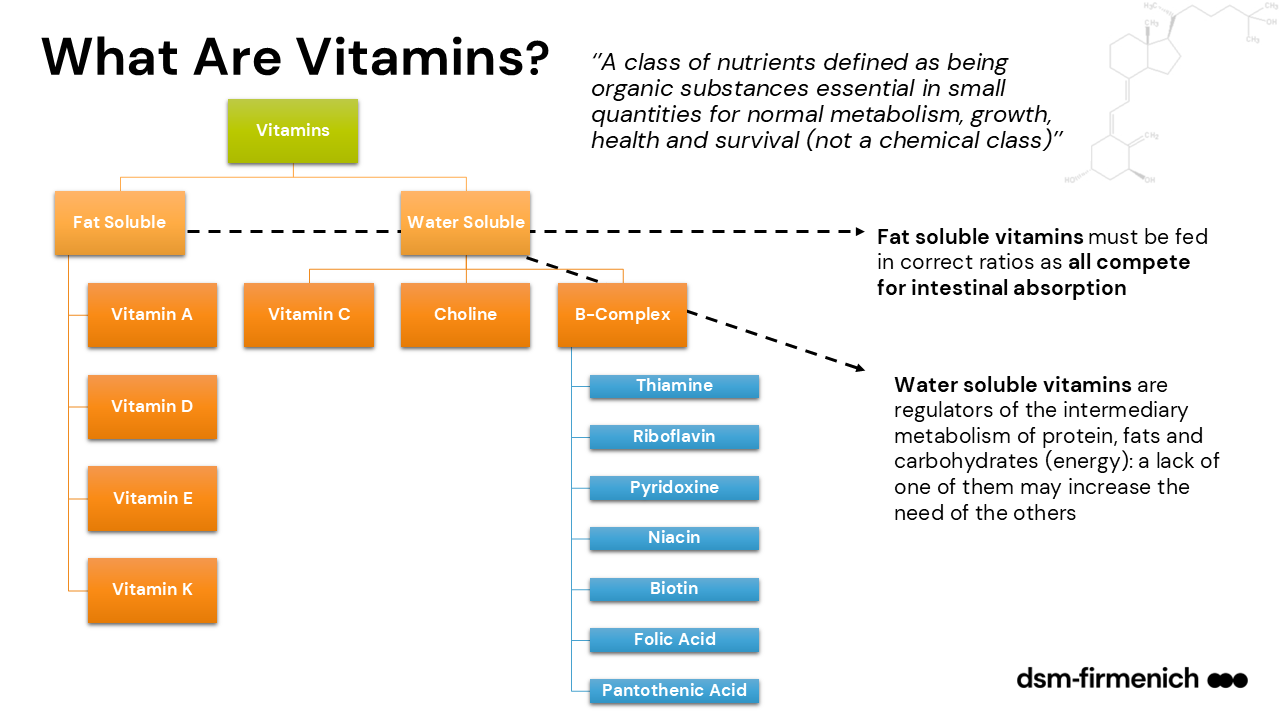 Figure 1
Figure 1
Vitamin requirements are affected by the physiological and production state of the animal, their health status and challenges, dry matter intake level and composition of the basal diet, nutritional status of the animal, seasonal fluctuations, geographical region, housing type and stress exposure. Feedlot cattle are subjected to numerous stressors impacting morbidity and mortality especially during the receiving period, and factors such as transport over long distances, processing, comingling, learning to eat from a bunk, changes in feed delivery, heat, cold and mud stress all contribute to oxidative stress at a cellular level, increasing the requirement for vitamin A and specifically, beta carotene (Shastak et al., 2023; NRC, 1996).
Vitamin A undergoes extensive microbial degradation in the rumen with 80% degradation in concentrate-based diets (devoid of natural occurring beta carotene from green plant material) and 20% degradation in forage-based diets (Rode et al., 1990, Calsimiglia et al., 2022). Beta carotene degradation in the rumen is estimated at 55% with 67% of retinol esters degraded. The animal obtains vitamin A either from plant feed ingredients in the form of beta carotene (the carotenoid pre-cursor to vitamin A) or as a retinol ester (synthetic esters of retinol such as vitamin A propionate or palmitate) supplemented in the diet. Both beta carotene and retinol esters, like all fat-soluble vitamins, rely on fat digestion for absorption and are absorbed mainly from the proximal jejunum with an intestinal absorption rate of 80-90% for retinol and 50-60% for beta carotene (Olsen, 1984; Bierer et al., 1995). As beta carotene and the retinol ester pass through the abomasum into the duodenum, pepsin aids the release of the beta carotenoids and retinol esters from the digesta and pancreatic retinol ester hydrolase enzyme hydrolyses the retinol esters into
retinol, the hydroxyl vitamer form. Absorption of the retinol and beta carotene across the brush border into enterocytes depends on solubilization within the bile salt micelles. At the low pH of the duodenum, bile facilitates fatty acids being transferred from the insoluble particulate phase/digesta into the micellar phase allowing passive diffusion of vitamins across the brush border into the intestinal mucosal cells. Beta carotene then undergo enzymatic cleavage into retinal (the aldehyde form of the vitamer) which will again be reduced to retinol and together with the retinol source from the original synthetic retinol ester supplement will be converted back to a retinol ester to be packaged together with any remaining uncleaved beta carotene into chylomicrons and lipoproteins, extruded into the lymphatic system and transported into the blood via the thoracic duct (Bonet et al., 2003). Retinal is important for vision and can be converted into all trans-retinoic acid (RA) through irreversible oxidation. In its retinoic acid form, vitamin A plays a major role in cell differentiation, hematopoiesis, embryonic development and cell growth and differentiation (Church, 1976). Retinoic acid is the main vitamer utilized in cattle (yellow fat animals) as the majority of beta carotene is not enzymatically cleaved, but incorporated into chylomicrons, stored in the liver and circulate in the blood as the lipoprotein fraction (Schweigert et al., 1987). Beta carotene is, however, converted within other tissues such as follicular structures (corpus luteum) to retinal and functions as a powerful antioxidant, complimentary to vitamin E’s antioxidant properties and synergistic to vitamin C and polyphenols (Leger et al., 2006). In the liver the retinol esters from the degraded chylomicron remnants are hydrolyzed into the hydroxyl form, retinol, where it is stored in the Kupfer cells in addition to storage within adipose tissue. Expression of retinol binding protein, required for solubilization and transport of retinol for esterification or oxidation are high in both liver and adipose tissue (Bonet et al., 2003, Engle et al., 2025). The liver is the primary storage organ and additionally 15-20% of retinoids are stored in adipose tissue (Tsutsumi et al., 1992).
Liver storage of vitamin A provides a 3-to-4-month supply in cattle depending on stage of production, stressors and level of nutrition (NASEM, 200, Rode et al., 1990). Vitamin A is released from the liver in its hydroxyl form (retinol) bound to retinol binding protein, critical for its mobilization from storage and aided by the transport protein, transthyretin to be transported to target tissues as a complex (Ikeda et al., 2005; Church, 1976). Turnover of liver vitamin A reserves are dependent on intake and function with feedlot cattle and dairy cows exhibiting rapid turnover estimated at 50% in 90-320 days.
Vitamin A Status
Retinol determination of liver tissue collected through biopsy or at harvest, serves as the gold standard to determine overall status, providing the animal is not deficient in vitamin A (OVN, dsm-firmenich, 2022). Due to tight homeostatic regulation of vitamin A , retinol concentration in serum or plasma is not the most accurate indication of animal status. Retinol is determined in blood and liver tissue using reversed-phase high-performance liquid chromatography (HPLC). A low or high retinol status can also be determined using dried blood spot technology (DBS) with a small blood sample collected under field conditions with limited infrastructure (Gannon et al., 2020; OVN, dsm-firmenich, 2022). Additionally, determining the beta carotene status of the animal provides an indirect measurement of vitamin A status considering beta carotene’s biological activity is in the order of 50% compared to retinol. Beta carotene can be assessed in plasma using HPLC coupled with visible spectrophotometry.
Recommendations for optimum serum and liver levels in finishing cattle as an indication of adequate vitamin A status are outdated and, in many cases, incomplete; however, Speer et al., 2022 describe an optimal serum retinol level of 225-325 ng/ml in calves, 300-800 ng/ml in cows and Wellmann et al.,2020 recorded a level of 225-500 in finishing cattle. Adequate liver status is described by Pulse, 1994 for the cow and calf only at 300-700 µg/g DM for cows and 100-350µg/g DM for calves.
The Reason for Supplementation
Vitamin A has proven to be essential to prevent night blindness (xeropthalmia) in cattle, a common symptom of vitamin A deficiency, as it supports vision through epithelial cell differentiation and retinal pigmentation. Vitamin A is also essential for immunity and reproduction (steroid synthesis), skin, epithelial and mucosal integrity (cell differentiation and gene transcription). Deficiency presents as a loss of appetite, poor nutrient absorption, impaired growth, reproduction effects in breeding stock due to decreased spermatogenesis and fetal resorption and losses, skin keratinization and increased risk of respiratory and intestinal infections (Kwada et al., 1996; NRC, 1996; Bonet et al., 2003).
Vitamin A is the one vitamin secreted in colostrum and milk at 100% of calf requirements (NASEM, 2021) at a level of 5,216 IU/l. Vitamin A supplementation of the gestational cow is thus important to ensure adequate status in the calf as demonstrated by Speer et al., 2023 recommending supplementing cows with a minimum of 93,000 IU/d for 165 days to elevate liver status of calves into the adequacy range (100-350 µg/kg DM). Supplementation, specifically in the form of beta carotene to breeding cows, is supported by research in dairy cows relating a higher level of beta carotene status to a reduction in pregnancy loss, increased breeding success and conception rates, less days open and elevated vitamin A status of the calf at birth (Madureira et al., 2020, Chorfi et al., 2010, Prom 2016). The calf loses its
elevated beta carotene status between day 7 and 60 and supplementation through the milk replacer, calf starter and later grower feed is recommended to maintain adequate vitamin A status.
Calves coming off pasture during the winter months or conditions of drought (marked by lower beta carotene content of plant forage) are more likely to have a lower vitamin A status which upon arrival at the feedyard would put them at a disadvantage, eroding the typical 3– 4-month supply of vitamin A reserves in the liver (Rode et al., 1990).
Animals with adequate vitamin A status have been shown to have a reduced level of oxidative stress resulting from stressors such as heat stress (Shastak et al., 2023; Kucuk et al., 2003; Hui et al., 2020; Shi et al., 2018).
Not discounting the important role of vitamin A supplementation to support the abovementioned functions, vitamin A restriction has also been widely investigated specifically in terms of its potential effects on marbling (intramuscular fat deposition) and growth in typical U.S. cattle breeds. Adipose tissue has access to two sources of retinol: Retinol esters hydrolyzed by lipoprotein lipase in the postprandial chylomicrons (Blaner et al., 1994) and retinol absorbed from the bloodstream, complexed to RBP with transthyretin and or albumin which when bound to cellular binding protein delivers retinol to target cells (Kurlandsky et al., 1995). Retinoic acid, retinal, retinol and beta carotene were recognized as inhibitors of terminal adipocyte differentiation mainly in research conducted with Hanwoo and Japanese Black Cattle (typically fed in confinement from birth to 30 months of age (Nade et al., 2003; Murray and Russel, 1980; Kawada et al., 1990; Hida et al., 1998; Ohyama et al., 1998 ; Kawada et al. 2000); however, the threshold level of retinol vitamers required for inhibition has not been identified (Xue et al., 10996). In an excellent review on the subject by Engle et al., 2025, the authors conclude that the effects of restrictive use of vitamin A supplementation are inconsistent and do not recommend deviating from NASEM recommendations for vitamin A supplementation. Research conducted by Bryant et al. in 2010 with typical black-yearling crossbred steers suggested vitamin A supplementation up to twice NASEM recommendation does not affect performance, marbling or lipogenic enzyme activity in adipose tissue. These findings agree with Pyatt et al., 2005; Gerocica-Buenfill et al., 2007c; Pickworth et al., 2012; Ward et al, 2012 and Gibb et al., 2011 reporting no impact of vitamin A restriction on marbling score or carcass grade.
Supplementation Guidelines
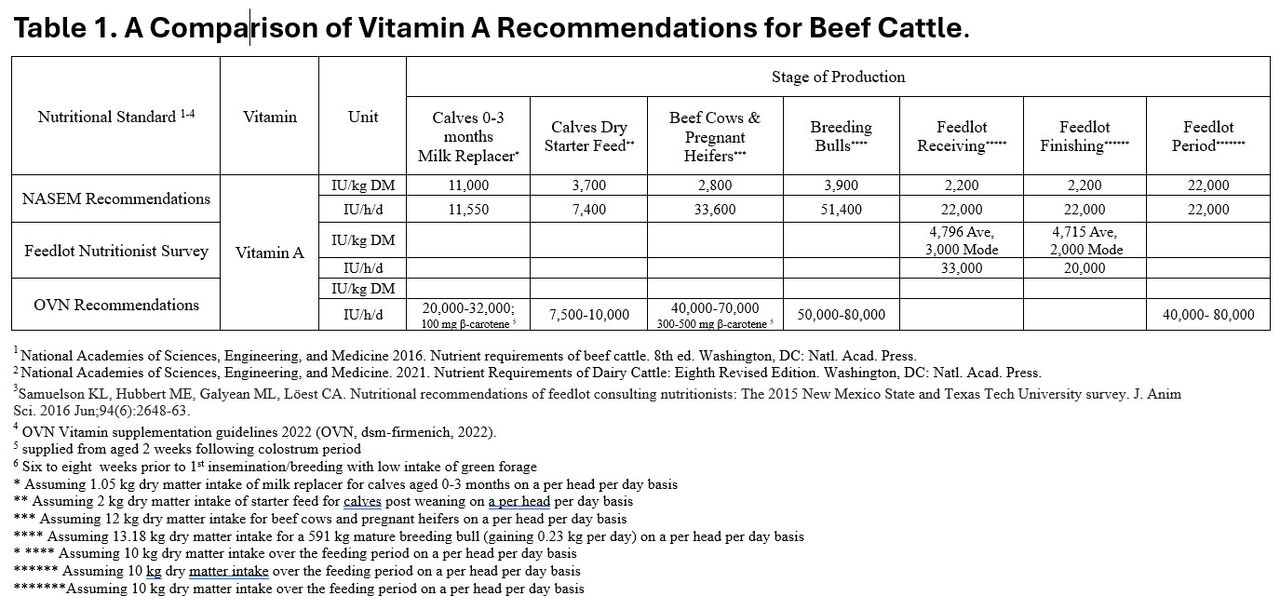
Vitamin A recommendations for finishing cattle varies between 22,000 and 80,000 IU/h/d (NASEM 2000; dsm-firmenich’s OVN, 2022 recommendations) compared with the Feedlot Consulting Nutritionists Survey reflecting industry practice aligns closely with NASEM recommendations (Table 1). Current research suggests these levels are sufficient to maximize health and productivity.
Following vitamin K, vitamin A is the most reactive vitamin with the least stability in vitamin premix mixtures and feeds and hence requires stabilization (Shurson et al., 1996).
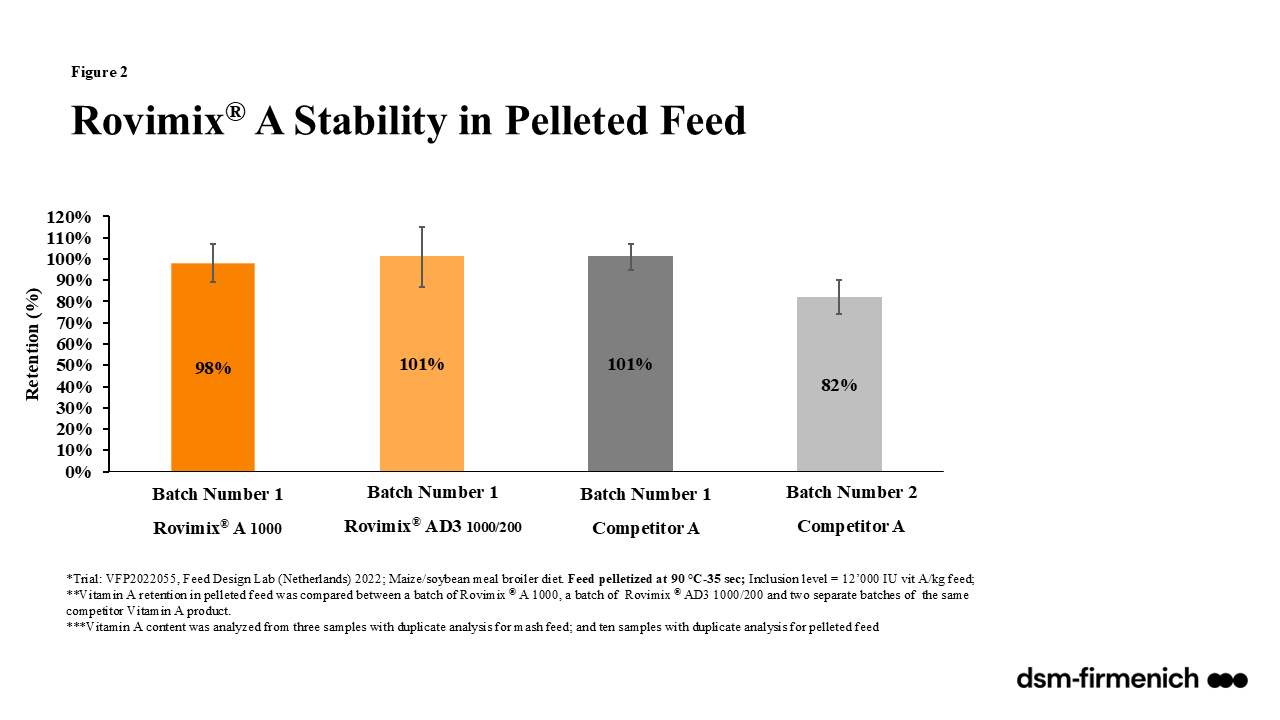
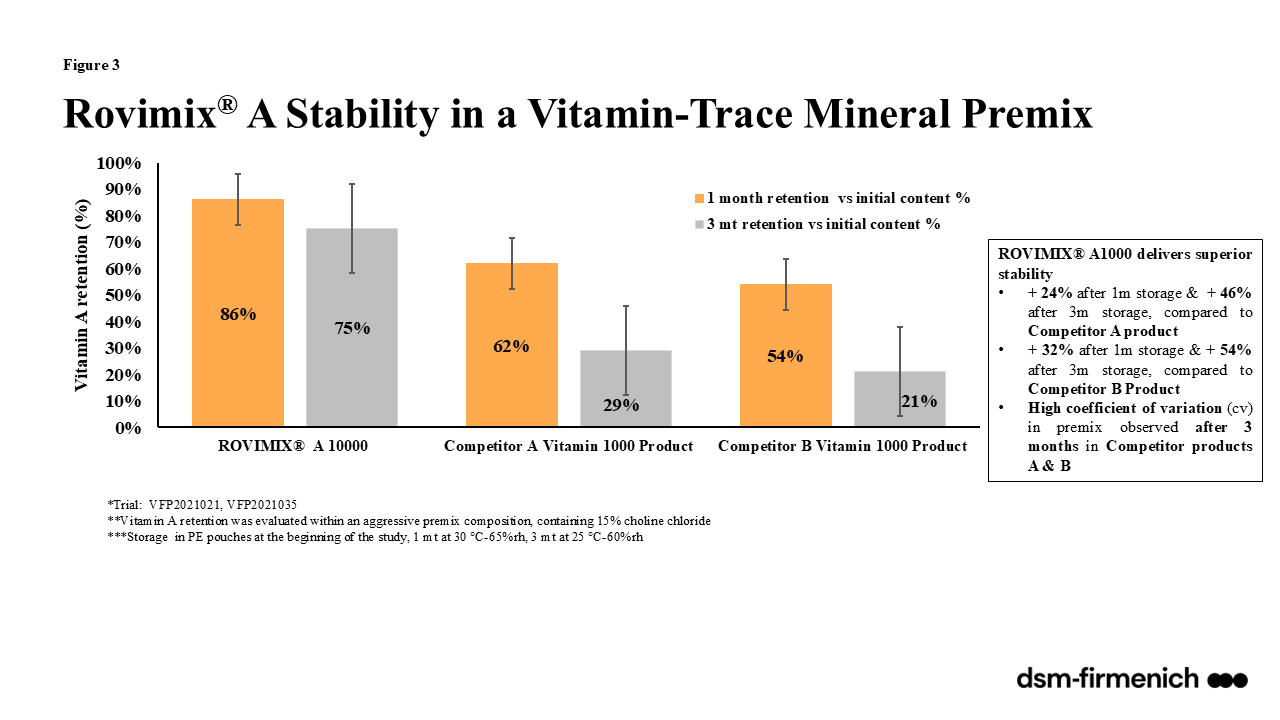
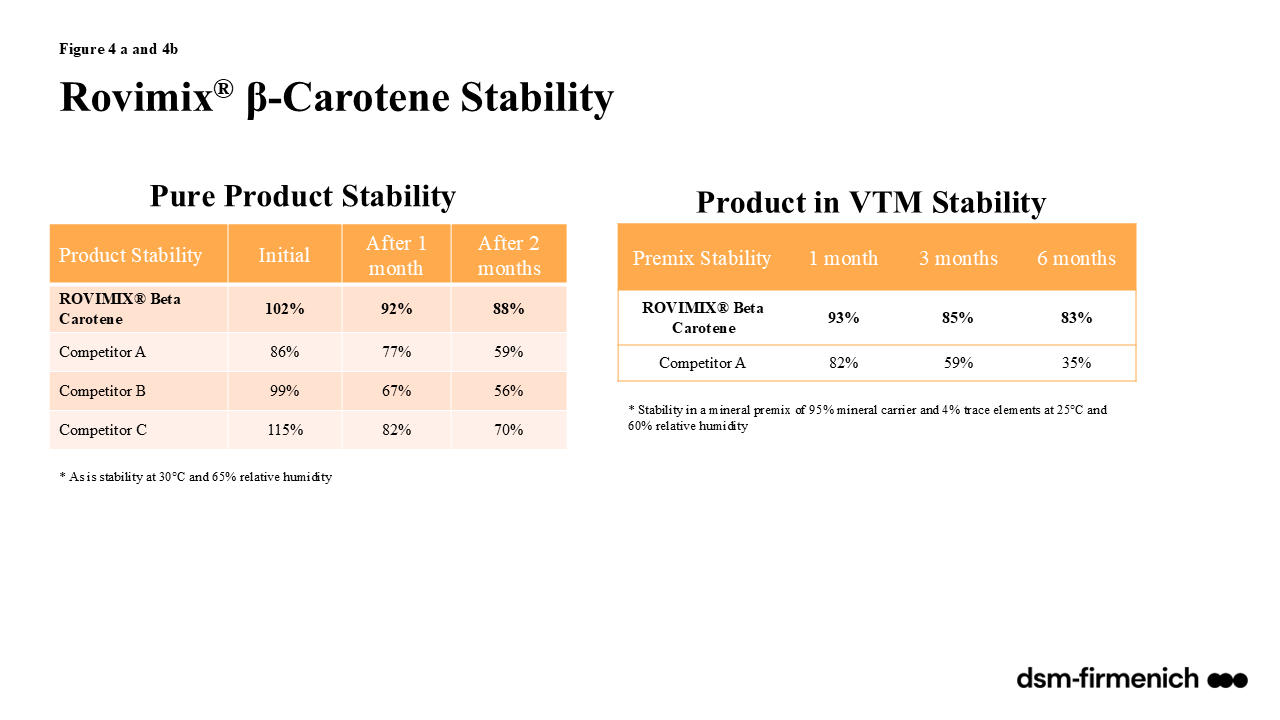
Vitamin A can be supplemented in the diet as a combination of synthetic esters of retinol and propionate and/or palmitate. Rovimix® A 1,000 (dsm-firmenich) provides a stabilized form of vitamin A in the form of vitamin A acetate, also described as retinyl acetate and all-trans vitamin A acetate. Rovimix® β-carotene 10% (dsm-firmenich) provides beta carotene stabilized with ascorbyl palmitate amongst other antioxidants providing stability for inclusion in all feeds. Numerous evaluations comparing the stability of Rovimix® Vitamin A and Rovimix® β-carotene amongst competitor products proved Rovimix® Vitamin A to be superior with 98-101% retention across pelleted feeds (90ºC for 35 seconds) (Figure 2) and 86% and 75% retention one and three months, respectively, following inclusion in an aggressive, choline chloride containing vitamin trace mineral mix (Figure 3). Rovimix® β-carotene 10% proved superior stability of the pure product 92% retention after one month and 88% after two months compared with competitors ranging between 67% at one month and 56% at two months (Figure 4a). Rovimix® β-carotene 10% included in a vitamin trace mineral mix had 83% retention, six months following inclusion compared with 35% retention of a competitor product (Figure 4b).
Supplementation of vitamin A in the form of dispersible liquid concentrates is an alternative, cost-effective option for vitamin A supplementation in cattle. Rovimix® Dispersible Liquid Concentrates (dsm-firmenich) are colloidal dispersions of fat-soluble vitamins mixed with food-grade emulsifiers and solvents. Vitamin A provided as a combination of vitamin A propionate and palmitate within the dispersible liquid concentrate stays in suspension when mixed into liquid feed supplements with no separation at high temperatures or gelatinization at low temperatures. Being dispersible and miscible with high viscosity liquids such as molasses, this application of vitamin A improves ease of use and is suited for addition to free-choice liquid feeds, concentrated liquid feed supplements and use in micro-dosing machines.
In reviewing vitamin A supplementation recommendations compared with industry practice, metabolism and function in cattle, considering the rate of ruminal and in feed degradation, efficiency of absorption, storage in the liver and adipose tissue in addition to what is considered an optimal or adequate range of vitamin A in the liver and serum as indication of animal status, vitamin A supplementation proves to be essential. Supplementation is advised to mitigate common deficiency symptoms such as night blindness in cattle but more importantly to optimize health through increased cell differentiation, reducing the risk of respiratory and intestinal infections, supporting intake and nutrient absorption in addition to enhanced reproductive effects in breeding stock.
The profitable finishing of feedlot cattle starts with the gestating cow. Reproductive efficiency of the cow is improved by vitamin A and specifically beta carotene supplementation (Madureira et al., 2020, Chrofi et al., 20101, Prom,2016; NRC, 1996, Speer et al., 2023). Vitamin A status of the newborn calf is influenced by that of the dam (NRC, 1996. Speer et al., 2023) with supplementation for at least the last 165 days recommended to elevate the vitamin A status of the calf into the adequacy range, providing colostrum and whole milk sufficient in vitamin A content to meet the calf’s requirements. However, vitamin A supplementation of milk replacer and calf starter and grower feed is advised to maintain vitamin A status and sustain the desired growth rate of the calf. Vitamin A supplementation across the entire chain of production should ensure that regardless of drought conditions, access to green fodder, various stressors such as transporting cattle for long distances, processing, comingling, heat stress (all contributing to oxidative stress) a calf enters the feedyard not in state of vitamin A depletion. A healthy receiving calf with adequate vitamin status provides the feedlot nutritionist with a good baseline to build upon and maximize health and growth response over the duration of the growing period.
Bierer, T. L., N. R. Merchen, and J. W. Erdman Jr. 1995. Comparative absorption and transport of five commoncarotenoids in preruminant calves. J. Nutr. 125, 1569–1577.
Blaner, W. S., Obunike, J. C., Kurlandsky, S. B., AI-Haideri M, Piantedosi, R., Deckelbaum, R. J., Goldberg, I. J. 1994. Lipoprotein lipase hydrolysis of retinyl ester. J. Biol. Chem 269:16559-16565.
Bonet, M. L., J. Ribot, and F. Felipe et al. Vitamin A and the regulation of fat reserves. CMLS, Cell. Mol. Life Sci. 60, 1311–1321 (2003). https://doi.org/10.1007/s00018-003-2290-x
Bryant, T. C., J. J. Wagner, J. D. Tatum, M. L. Galyean, R. V. Anthony, and T. E. Engle. 2010. Effect of dietary supplemental vitamin A concentration on performance, carcass merit, serum metabolites, and lipogenic enzyme activity in yearling beef steers. J. Anim. Sci. 2010. 88:1465–1478.
Calsimiglia, S. Oviedo-Rondon, E.O., Tamassia, L., Litta, G. and Hernandez, J. M. Optimum Vitamin Nutrition. OVN Vitamin supplementation guidelines 2022.
Chorfi, Y. 2010 «Impact of peripartum beta-carotenemia on subsequent reproductive performance in Holstein Dairy Cows (personal communication).
Church. D. C. The Ruminant Animal. Digestive physiology and nutrition of ruminants. 1976. Oregan State University. CME feeder cattle index. July 2025. Accessed July 31, 2025. Feeder Cattle Prices and Feeder Cattle Futures Prices - Barchart.com
Gannon, B. M., C. Jones, & S. Mehta (2020). Vitamin A Requirements in Pregnancy and Lactation. Current developments in nutrition, 4(10), nzaa142. https://doi.org/10.1093/cdn/nzaa142
Gibb, D. J., Van Herk, F. H., Mir, P. S., Loerch, S. and McAllister, T. A. 2011. Removal of supplemental vitamin A from barley-based diets improves marbling in feedlot heifers. Can. J. Anim. Sci. 91: 669674. T
Gorocica-Buenfil, M. A. F. L. Fluharty, T. Bohn, S. J. Schwartz, and S. C. Loerch. 2007c. Effect of low vitamin A diets with high-moisture or dry corn on marbling and adipose tissue fatty acid composition of beef steers. J. Anim. Sci. 85:3355–3366.
Hida, Y., T. Kawada, S. Kayahashi, T. Ishihara, and T. Fushiki. 1998. Counteraction of retinoic acid and 1,25-dihydroxyvitamin D3 on up-regulation of adipocyte differentiation with PPARγ ligand, an antidiabetic thiazolidinedione, in 3T3-L1 cells. Life Sci. 62:205.
Hui J, Li L, Li R, Wu M, Yang Y, Wang J, Fan Y, Zheng X. 2020. Effects of supplementation with β-carotene on the growth performance and intestinal mucosal barriers in layer-type cockerels. Anim Sci J. 91(1): doi:10.1111/asj.13344
Ikeda, 2005. The Roles of Vitamin A for Cytoplasmic Maturation of Bovine Oocytes. Journal of Reproduction and Development, Vol. 51, No. 1, 2005 jrd51_1_23(06)-3.fm.fm (jst.go.jp)
Kawada, T., N. Aoki, Y. Kamei, K. Maeshige, S. Nishiu, and E. Sugimoto. 1990. Comparative investigation of vitamins and their analogues on terminal differentiation, from preadipocytes to adipocytes, of 3T3-L1 cells. Comp. Biochem. Physiol. 96A:323.
Kawada, T., Y. Kamei, and E. Sugimoto. 1996. The possibility of active forms of vitamins A and D as suppressors on adipocyte development via ligand-dependent transcriptional regulators. Int. J. Obes. Relat. Metab. Disord. 20(Suppl. 3):S52-57.
Kawada, T., Y. Kamei, A. Fujita, Y. Hida, N. Takahashi, E. Sugimoto, and T. Fushiki. 2000. Carotenoids and retinoids as suppressors on adipocyte differentiation via nuclear receptors. Biofactors 13:103-109.Kucuk O, Sahin N, Sahin K. 2003. Supplemental zinc and vitamin A can alleviate negative effects of heat stress in broiler chickens. Biol Trace Elem Res. 94:225–236. doi:10.1385/BTER:94:3:225.
Kurlandsky, S. B., M. V. Gamble, R. Ramakrishnan, and W. S. Blaner. 1995. Plasma delivery of retinoic acid to tissues in the rat. J. Biol. Chem. 270:17850.
Léger, 2006 (in french). Anti-oxydants d’origine alimentaire : diversité, modes d’action anti-oxydante, interactions. OCL VOL. 13 N° 1 JANVIER-FE´VRIER 2006 Anti-oxydants d’origine alimentaire : diversité, modes d’action anti-oxydante, interactions | OCL - Oilseeds and fats, Crops and Lipids (ocl-journal.org)
Madureira, A. M. L., Pohler, K. G., Guida, T. G., Wagner, S. E., Cerri, R. L. A., & Vasconcelos, J. L. M. (2020). Association of concentrations of beta-carotene in plasma on pregnancy per artificial insemination and pregnancy loss in lactating Holstein cows. Theriogenology, 142, 216–221. https://doi.org/10.1016/j.theriogenology.2019.10.006
Murray, T., and T. R. Russell. 1980. Inhibition of adipose conversion of murine 3T3L1 cells by retinoic acid. J. Supramol. Struc. 14:255-266.Nade et al., 2003; National Research Council (NRC) (1996) Nutrient Requirements for Beef Cattle. 7th Edition, The National Academies Press, Washington DC.
NASEM. National Academies of Sciences, Engineering, and Medicine. 2016. Nutrient Requirements of Beef Cattle: Eighth Revised Edition. Washington, DC: The National Academies Press.
NASEM. National Academies of Sciences, Engineering, and Medicine. 2021. Nutrient Requirements of Dairy Cattle: Eighth Revised Edition. Washington, DC: The National Academies Press. https://doi.org/10.17226/25806.
Neville, B. W., C. S. Schauer, K. Karges, M. L. Gibson, M. M. Thompson, L. A. Kirschten, N. W. Dyer, P. T. Berg, and G. P. Lardy. 2010. Effect of thiamine concentration on animal health, feedlot performance, carcass characteristics, and ruminal hydrogen sulfide concentrations in lambs fed diets based on 60% distillers dried grains plus solubles. J. Anim. Sci. 88:2444–2455.Olson, J. A. (1988) Biological actions of carotenoids. J. Nutr. 119: 94–95Olkowski, A. A., S. R. Gooneratne, C. G. Rousseaux, and D. A. Christensen. 1992. Role of thiamine status in sulphur induced polyencephalomalacia in sheep. Res. Vet. Sci. 52:78–85.
Ohyama, M., K. Matsuda, S. Torii, T. Matsui, H. Yano, T. Kawada, and T. Ishihara. 1998. The interaction between vitamin A and thiazolidinedione on bovine adipocyte differentiation in primary culture. J. Anim. Sci. 76:61-65.
Pickworth, C. L., S. C. Loerch, and F. L. Fluharty. 2012. Restriction of vitamin A and D in beef cattle finishing diets on feedlot performance and adipose accretion. J. Anim. Sci. 90:1866-1878.
Prom, C. M. 2016. Supplementation of β-carotene during the close-up period on cows, colostrum and calves, MS Thesis. Department of Animal Sciences, University of Illinois at Urbana-Champaign (2016) http://hdl.handle.net/2142/93065, Accessed 26th Aug 2020
Pulse, R. 1994. Vitamin Levels in Animal Health: diagnostic data and bibliographies. Sherpa International.
Pyatt, N. A., and L. L. Berger. 2005. Review: Potential effects of vitamins A and D on marbling deposition in beef cattle. Prof. Anim. Sci. 21:174-181. 69
Rode, L. M., T. A. McAllister, and K. J. Cheng. 1990. Microbial degradation of vitamin A in rumen fluid from steers fed concentrate, hay or straw diets. Can. J. Anim. Sci. 70:227-233.
Samuelson, K. L., M. E. Hubbert, M. L. Galyean, and C. A. Löest. Nutritional recommendations of feedlot consulting nutritionists: The 2015 New Mexico State and Texas Tech University survey. J Anim Sci. 2016 Jun;94(6):2648-63. doi: 10.2527/jas.2016-0282. PMID: 27285940.
Schweigert, Florian J. et al. “Plasma and tissue concentrations of beta-carotene and vitamin A in rats fed beta-carotene in various fats of plant and animal origin.” Journal of environmental pathology, toxicology and oncology : official organ of the International Society for Environmental Toxicology and Cancer 19 1-2 (2000): 87-93.
Shastak, Y., Gordillo, A., and Pelletier, W. 2023. The relationship between vitamin A status and oxidative stress in animal production. Journal of Applied Animal Research, 51(1), 546–553. https://doi.org/10.1080/09712119.2023.2239319
Shi, H., Yan, S., Guo, Y., Shi, B., and Guo, X. 2018. The pre-protective effect of vitamin A on LPS-induced oxidative stress of bovine mammary epithelial cells. Italian Journal of Animal Science, 17(4), 959–966. https://doi.org/10.1080/1828051X.2018.1453757
Shurson, J., T. Salzer,, and D. Koehler. 1996. Metal-specific amino acid complexes, inorganic trace minerals effect on vitamin stability examined Feedstuffs, 68 (1996), pp. 13-23.
Smith, R. M. Cobalt. In: Mertz W, editor. Trace elements in human and animal nutrition. New York: Academic Press; 1987. p. 143–83.
Speer, H., H. Freetly, and M. E. Drewnoski. 2022. 11 Evaluating Relationships Between Plasma and Liver Retinol Concentrations in the Beef Cow and Calf, Journal of Animal Science, Volume 100, Issue Supplement_4, November 2022.
Speer, H., K. Wilke, and M. E. Drewnoski. 2023. 53 Effect of Vitamin A Supplementation During Gestation on Vitamin A Status of Beef Cows and Their Calves Managed in the Drylot, Journal of Animal Science, Volume 101, Issue Supplement_2, November 2023, Pages 49 50, https://doi.org/10.1093/jas/skad341.054
Tsutsumi, C., M. Okuno, L. Tannous, R. Piantedosi, M. Allan, D. S. Goodman, and W. S. Blaner. 1992. Retinoids and retinoid-binding protein expression in rat adipocytes. J. Biol. Chem. 267:1805.
Ward, E. H., and H. H. Patterson. 2004. Effects of thiamin supplementation on performance and health of growing steers consuming high sulfate water. Proc. West Sec. Am. Soc. Anim. Sci. 55:375–378.
Ward, A. K., J. J. McKinnon, S. Hendrick, and F. C. Buchanan. 2012. The impact of vitamin A restriction and ADH1C genotype on marbling in feedlot steers. J. Anim. Sci. 90:2476-2483.
Wellmann, K. B., J. Kim, P. M. Urso, Z. K. Smith, and B. J. Johnson. Evaluation of the dietary vitamin A requirement of finishing steers via systematic depletion and repletion, and its effects on performance and carcass characteristics. J Anim Sci. 2020 Sep 1;98(9):skaa266. doi: 10.1093/jas/skaa266. PMID: 32812033; PMCID: PMC7505414.
Xue, J. C., E. J. Schwarz, A. Chawla, and M. A. Lazar. 1996. Distinct stages in adipogenesis revealed by retinoid inhibition of differentiation after induction of PPARγ. Mol. Cell. Biol. 16:1567-1575.
18 August 2025
20 Jul 2026
Account Manager, Michelle Lutz, provides an overview of the benefits of vitamin A and beta-carotene for reproductive performance in cattle.

20 Jul 2026
Modern feed industry quality management has shifted from relying solely on finished-product testing to a proactive, risk-based approach focused on prevention. Manufacturers build quality into every stage of production through supplier qualification, ingredient verification, process controls, equipment maintenance, and data-driven monitoring. Industry standards such as FAMI-QS, ISO 9001, FSMA, and Canadian Feeds Regulations support this preventive philosophy. By identifying and addressing potential issues early, companies improve product consistency, regulatory compliance, operational efficiency, and traceability. Ultimately, effective quality systems reduce variability, strengthen customer confidence, and ensure safe, reliable premixes are produced consistently.
15 Jun 2026
The vitamin market enters Q3 2026 with a firm but stabilizing, cost-driven outlook, supported by geopolitical tensions, elevated energy costs, and ongoing supply constraints. This follows a broad price correction observed in June across several key vitamins, though downside risk remains limited. While some segments are relatively balanced, others remain tight due to disruptions and disciplined supplier behavior. Buyer strategies vary, though fewer participants remain in a wait-and-see mode, reflecting expectations of prolonged market tightness and uncertain normalization timing. Demand trends are uneven across regions, contributing to varied market positioning. Overall, conditions are expected to remain firm, with selective tightness, continued volatility, and a strong focus on supply risk management amid limited forward visibility.