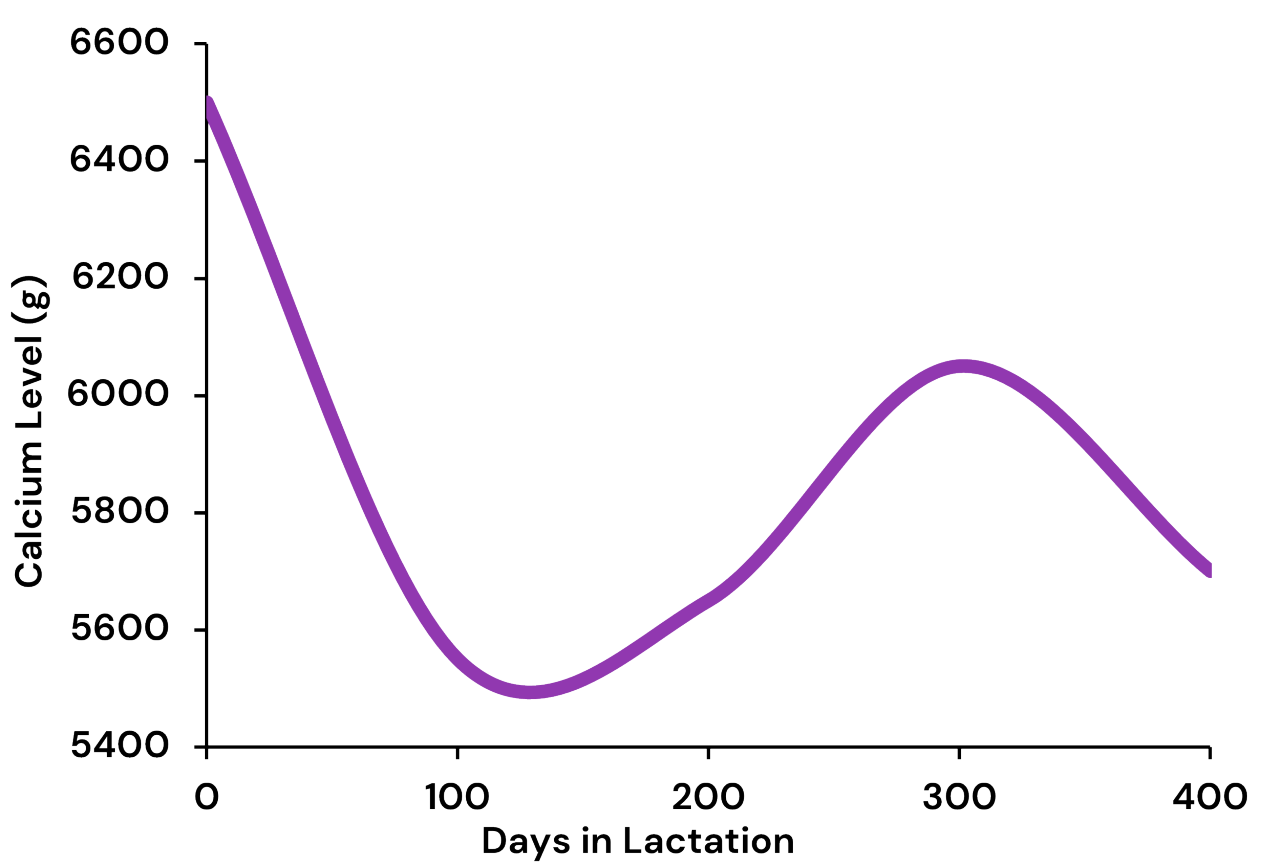
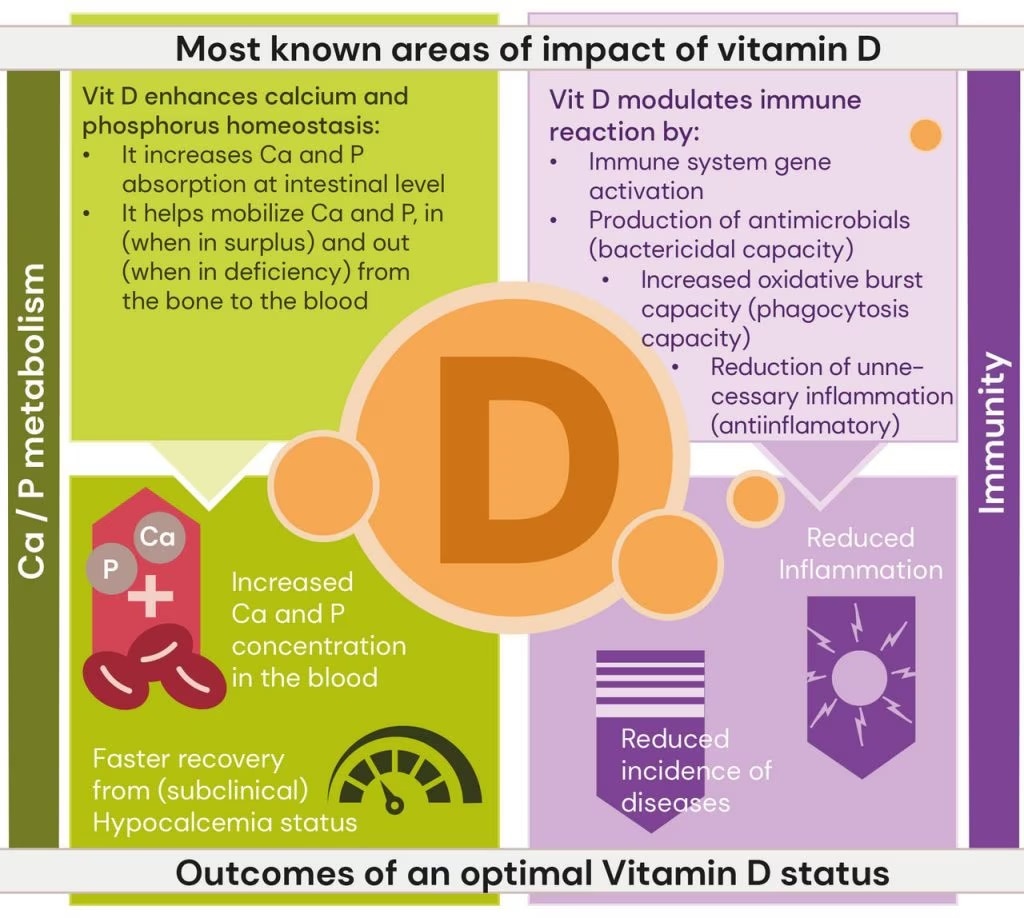
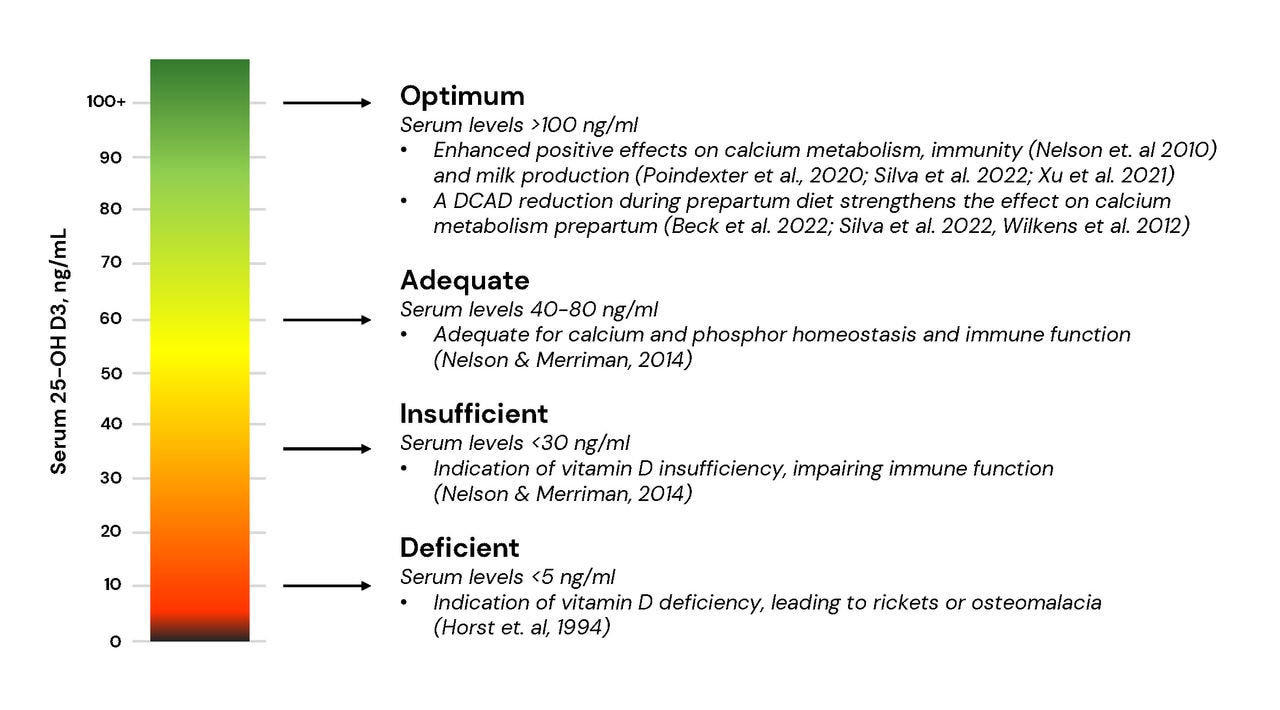
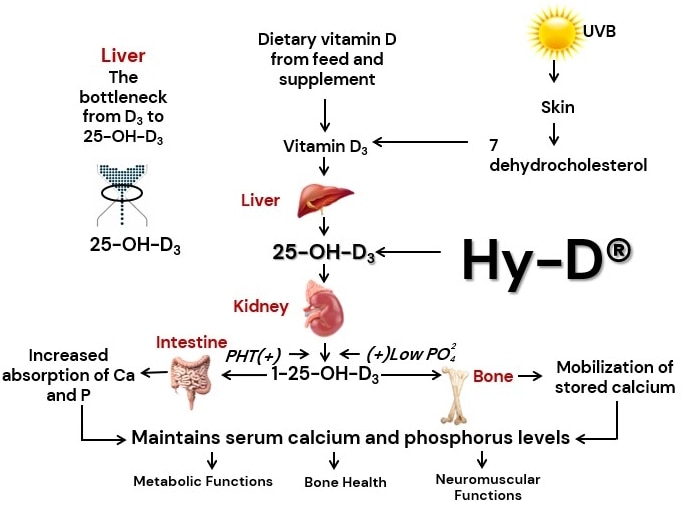
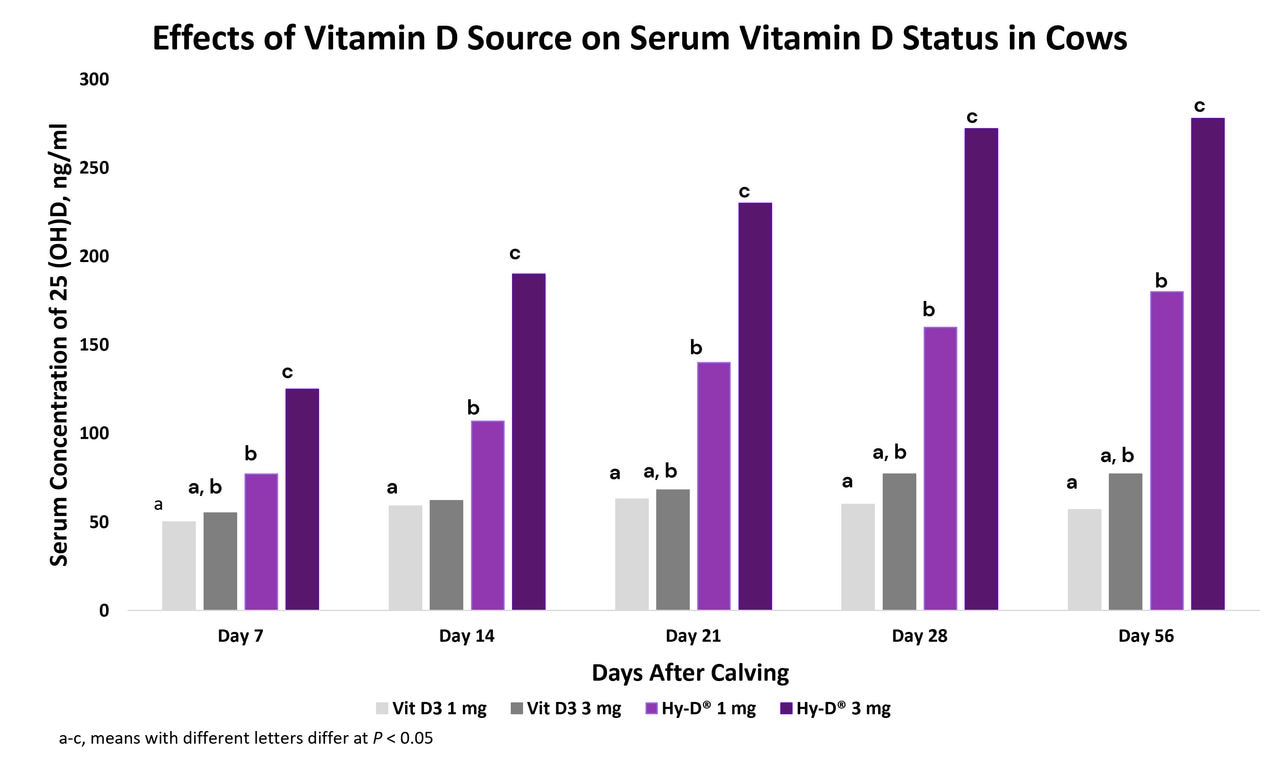
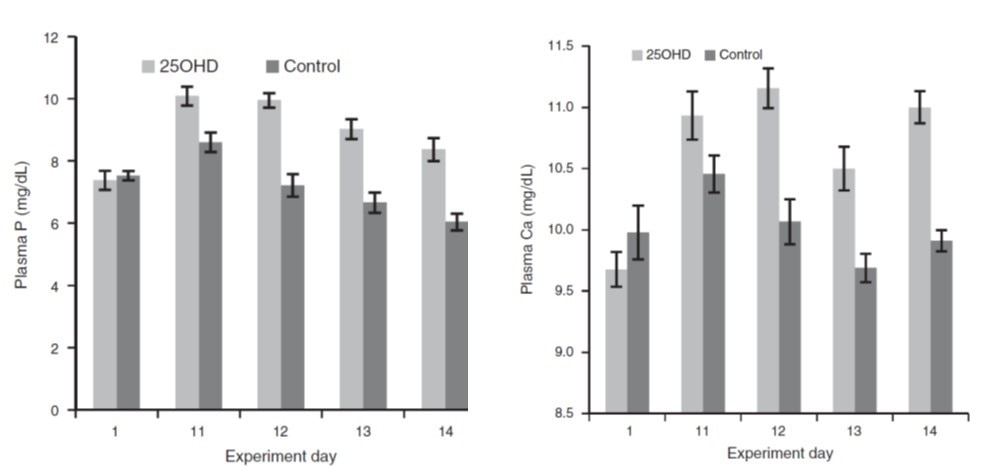
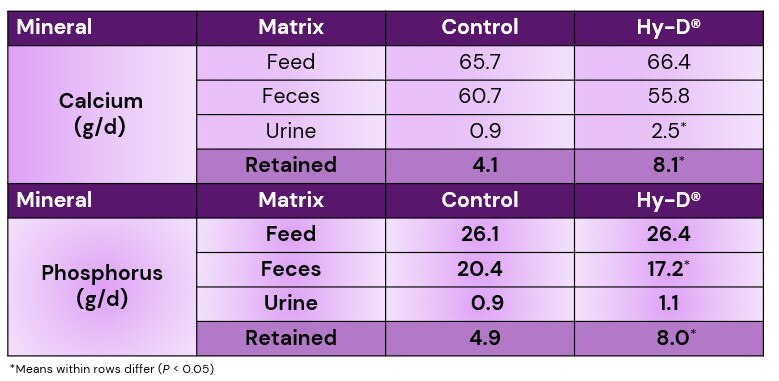
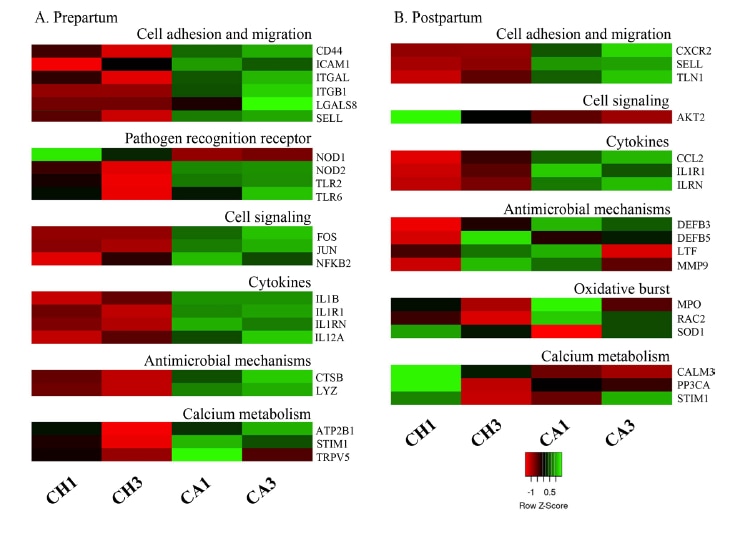
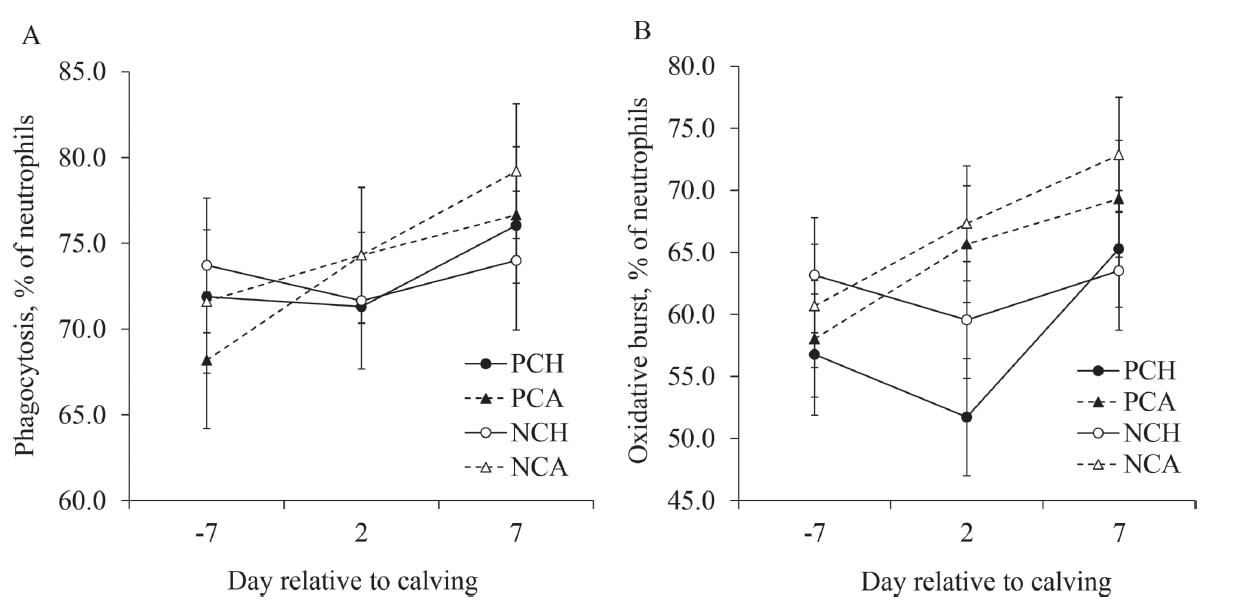
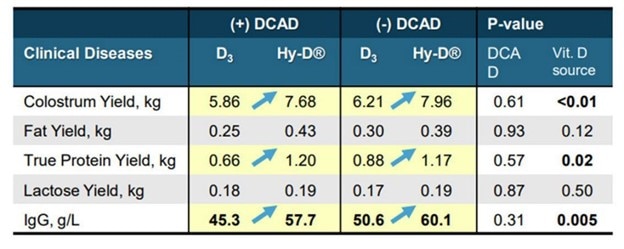
Beck et al. 2022. Effect of vitamin D source and dietary cation–anion difference in peripartum dairy cows on calcium homeostasis and milk production. Translational Animal Science, 6(1).
Goff, J. P., R. L. Horst, and E. T. Littledike. 1982. Effect of the ma[1]ternal vitamin D status at parturition on the vitamin D status of the neonatal calf. J. Nutr. 112:1387–1393.
Gombart, A.F. 2009. The vitamin D–antimicrobial peptide pathway and its role in protection against infection. Future Microbiology, 4: 1151.
Horst et al. 1994. Calcium and vitamin D metabolism in the dairy cow. Journal of Dairy Science, 77. 1936-1951
Kronfeld, D.S. 1976. The potential importance of the proportions of glucogenic, lipogenic and aminogenic nutrients in regard to the health and productivity of dairy cows. Advanced Animal Nutrition and Animal Physiology, 7. 7.
Martinez et al. 2018a. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Lactation performance and energy metabolism. Journal of Dairy Science, 101. 2544-2562.
Martinez et al. 2018b. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Health and reproductive responses. Journal of Dairy Science, 101. 2563-2578.
McGrath J. J., Savage D. B., Nolan J. V., Elliott R. 2012. Phosphorus and calcium retention in steers fed a roughage diet is influenced by dietary 25OH-vitamin D. Animal Production Science, 52(7). 636-640.
McGrath, J., Savage, D., Nolan, J. and Elliott, R. 2012a. Phosphorus and calcium retention in steers fed a roughage diet is influenced by dietary 25OH-vitamin D. Journal of Animal Production Science 52 (6-7), 636-640.
McGrath et al. 2015. The role and potential advantages of vitamin D metabolites in maintaining calcium status in high-producing dairy herds. Animal Production Science, 55(9). 1081-1089.
Nelson & Merriman. 2014. Vitamin D Metabolism in Dairy Cattle and Implications for Dietary Requirements. Conference Paper. Florida Ruminant Nutrition Symposium.
Nelson et al. 2010. Modulation of the bovine innate immune response by production of 1a, 25-dihydroxyvitamin D3 in bovine monocytes. Journal of Dairy Science, 93(3), pp.1040-1049.
Nelson et al. 2016. Vitamin D status of dairy cattle: Outcomes of current practices in the dairy industry. J. Dairy Sci. 99:10150–10160
Poindexter et al. 2020. Feeding supplemental 25-hydroxyvitamin D3 increases serum mineral concentrations and alters mammary immunity of lactating dairy cows. Journal of Dairy Science, 103(1). 805-822.
Silva et al. 2022. Effects of feeding 25-hydroxyvitamin D3 with an acidogenic diet during the prepartum period in dairy cows: Mineral metabolism, energy balance, and lactation performance of Holstein dairy cows. Journal of Dairy Science, 105(7). 5796-5812.
Standing Committee on Agriculture (SCA). 1990. Feeding Standards for Australian Livestock – Ruminants. CSIRO Publications, Melbourne. 1-17.
Vieira-Neto et al. 2021. Effect of source and amount of vitamin D on function and mRNA expression in immune cells in dairy cows. Journal of Dairy Science, 104(10). 10796-10811.
Weiss, W. P., E. Azem, W. Steinberg, and T. A. Reinhardt. 2015. Ef[1]fect of feeding 25-hydroxyvitamin D3 with a negative cation-anion difference diet on calcium and vitamin D status of periparturient cows and their calves. J. Dairy Sci. 98:5588–5600.
Wilkens et al. 2012. Influence of the combination of 25-hydroxyvitamin D3 and a diet negative in cation-anion difference on peripartal calcium homeostasis of dairy cows. Journal of Dairy Science, 95 (2012). 151-164.
Xu et al. 2021. Effects of 25-Hydroxyvitamin D3 and Oral Calcium Bolus on Lactation Performance, Ca Homeostasis, and Health of Multiparous Dairy Cows. Animals, 11(6). 1576.