Animal Nutrition & Health

The word ‘unprecedented’ is used liberally and often inappropriately. However, the current macro-economic and global political landscape is currently going through genuinely unprecedented flux. Economic recovery post-COVID has driven strong inflationary pressures that are compounded by labor shortages and supply chain fragility. Global energy prices are extremely high, and many food staples are increasingly unaffordable for consumers. The conflict in Ukraine has further exacerbated commodity supply, especially for wheat, sunflower meal, oil and corn, and this is putting upward pressure on prices of these, and alternative, raw materials such as by-products and inorganic phosphates. As poultry and swine producers operate under tight margins and given that feed represents around 70% of total production cost, headwind associated with high ingredient prices has a catastrophic effect on profitability and food chain security.
“Given that feed represents around 70% of total production cost, headwind associated with high ingredient prices has a catastrophic effect on profitability and food chain security.”
Exogenous feed enzymes such as carbohydrase, protease and phytase have been used to enhance digestibility of cereals, grain, legumes and oilseed meals for several decades. Xylanases generate substantial increases in energy and amino acid retention via hydrolysis of hemicellulose. Hydrolysis end products (xylo-oligosaccharides) are fermented by the hind gut microbiome which generates improvements in net energy and amino acid digestibility, in part via hormonal regulation of gastric digestion. Proteases solubilize dietary protein that is otherwise recalcitrant to digestion and alleviates negative pressure on endogenous protein synthesis and loss from the gut. These effects generate measurable increases in amino acid and energy digestibility. Finally, phytases hydrolyse phytic acid, generating free phosphate and myoinositol while concurrently reducing the antinutritional effect of phytate as an intestinal irritant. This generates a substantial increase in digestibility of minerals, amino acids and energy and assists the host with productive partitioning of nutrients.
Reducing feed cost with exogenous enzymes
Each class of feed enzyme carries a unique nutritional matrix that can be deployed during least cost feed formulation to displace more expensive sources of minerals, amino acids and energy. When used in combination the feed cost savings are appreciable (>US$30/metric ton of feed) and these feed cost savings can be inflated by simultaneously relaxing constraints on by-product maximums. dsm-firmenich and Novozymes have worked in a cooperative alliance for more than 20 years and have brought several disruptive enzyme technologies to the market during this time (Table). The combined use of these enzymes can generate substantial nutrient release values that mitigate feed cost pressure. Our recommendations also consider the lack of full additivity on nutritional matrices of adjacent enzymes and are all fully supported by peer-reviewed meta-analyses and independent studies with opinion leading professors.
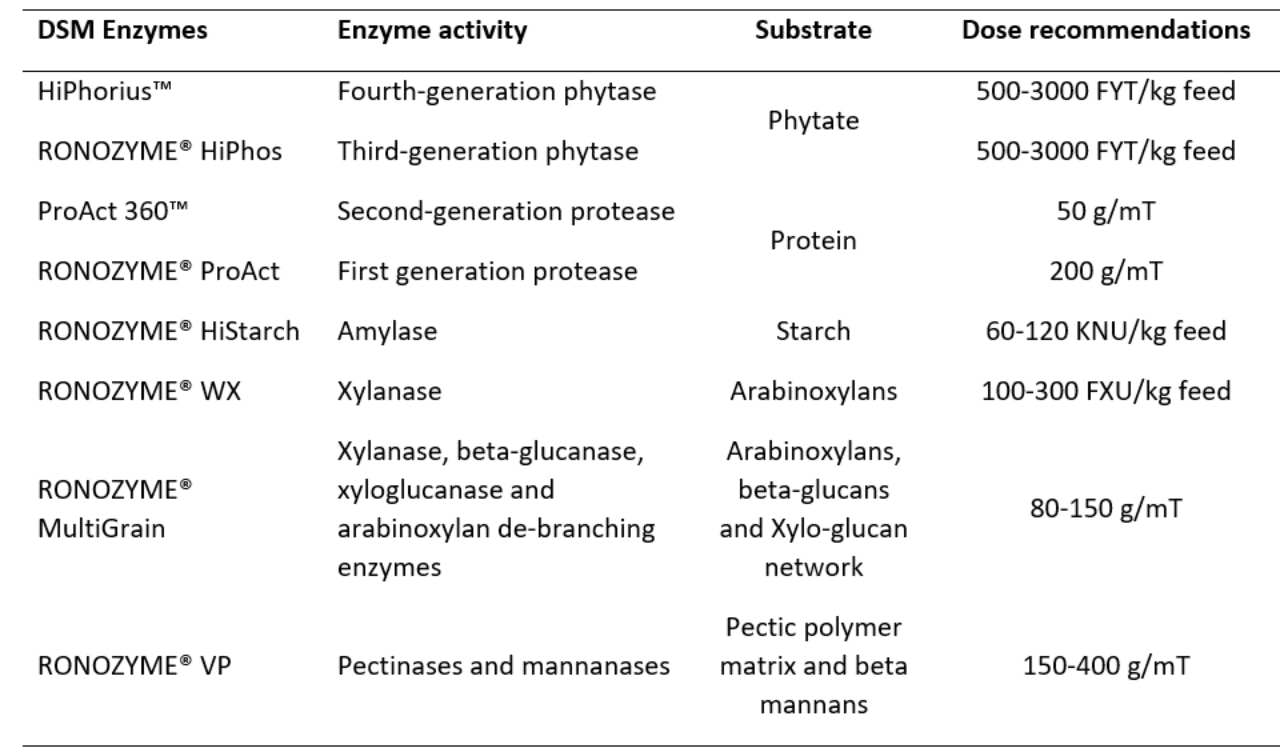
Table. dsm-firmenich Enzyme portfolio, enzyme activities, target substrates and dose recommendations
Optimizing value of feed enzymes
1. Substrate surveillance. Unlike many feed additives (e.g., essential oils or organic acids), feed enzymes hydrolyze specific and measurable substrates in raw materials. Near infrared spectroscopy (NIR) calibrations now exist for most of these substrates including non-starch polysaccharide fractions, protein solubility, cereal quality and phytic acid. As feed cost pressure increases motivation to proportionately elevate matrices for feed enzymes, the importance of mapping the substrate landscape in feed also increases so that animal performance does not suffer. There are several strategies that can be employed. For example, if a given diet is analytically confirmed to have relatively high phytic acid concentrations (>0.25% phytate phosphorus), higher doses of phytase and more aggressive displacement of inorganic phosphate can be accommodated. Increasing access to phytate to further optimize its complete removal can also be accomplished by use of adjacent additives such as accessory enzymes (especially protease), organic acids, vitamin D3 and 25-OH D3, management of drinking water pH, etc. A similar approach can be taken to align energy matrices with cereal quality. Corn or wheat with a high fiber concentration or a protein/starch matrix that is mechanically resistant to solubilization in the intestine may have a relatively low metabolizable energy concentration and in such cases the energy assumptions for carbohydrase can be inflated.
2. Cheaper feed ingredients. Relaxing maximum constraints on high fiber by-products and lower cost, locally sourced raw materials can also result in substantial reductions in feed cost but may come at the expense of FCR if this is not managed strategically. Higher fiber ingredients such as wheat bran, rice bran, palm kernel meal and distillery by-products contain fiber at concentrations sufficient to interfere with feed intake and to slough the mucosa of the intestinal tract. Thanks to NIR technology, these changes in dietary fiber profile can be quantified and mapped systematically to allow tactical enzyme selection. Importantly, high fiber by-products often contain appreciable phytate concentrations and this may be further considered in phytase dose optimization discussions.
3. Leveraging the microbiome. Use of mono-component carbohydrases and blends of complementary carbohydrases such as xylanases, glucanases, pectinases and amylase result in substantial increases in metabolizable, digestible and net energy density of feed ingredients and improve gut health via changes in fermentation profiles in the hind gut microbiota. These changes in the enteric microbiome are also cumulative and enable the host to build increasing tolerance to fiber ingestion. Importantly, to generate persistent beneficial effects, enzymes must be included from hatch to slaughter. Adding and removing enzymes in different dietary phases generates effects that are less predictable and more variable. Beyond energy, improvements in amino acid digestibility also occur when carbohydrases are utilised, and this can have further feed cost impact or generate reductions in FCR.
4. Low protein diets. Exogenous protease, supported by adjacent enzymes such as phytase and carbohydrase and strategic use of crystalline amino acids, allow animal performance to be sustained with radically reduced dietary crude protein concentrations. This has a substantial effect on diet cost and brings more latitude in formulation approaches e.g., reduced reliance on imported protein meals. Importantly, biochemical flow of protein and non-protein nitrogen to the environment is significantly reduced when dietary protein concentration is lowered e.g., for every 1% drop in dietary crude protein, ammonia nitrogen output is reduced by 10%. These changes not only influence feed cost but also improve animal health and welfare, litter quality, carcass quality and environmental sustainability. Upward ‘drift’ in FCR when radically low protein diets are fed may be mitigated via additional supply of digestible phosphate, combined with strategic use of carbohydrase and protease, to increase dietary protein retention and reduce endogenous amino acid loss. It is critical that an accurate amino acid matrix is applied as generic or ‘flat’ matrices for amino acids that do not consider the specific mode of action of the enzymes for different feed ingredients will result in unpredictable performance outcomes.
5. Precision nutrition. Finally, a dialogue with a local feed enzyme technical specialist is recommended because application of enzyme matrices, admixture selection and dosing should be done, whenever practical, with prior knowledge of the basal diet and live production targets. Tailored recommendations can be delivered that best suit the specific goals of the nutritionist, diet composition and desired formulation approaches.
Feed Cost Savings
When new feed enzyme products are introduced to the market, they typically carry a recommended nutrient release profile or ‘nutritional matrix’. These matrices are constructed during product development and are usually an aggregation of effects detected during multiple controlled experiments in university settings. However, in practice these matrices are only partially applied by commercial nutritionists and in some cases feed enzymes are used ‘on top’ of formulation where a small increase in feed cost is tolerated in return for a deferred improvement in FCR. Optimal value creation strategies for feed enzymes varies depending on meat and egg prices, the cost of feed ingredients and the shadow prices of nutrients in formulation. Today, as raw material prices are high and expected to remain so for the foreseeable future, a reassessment of nutrient matrices for feed enzymes is justified.
1. Elevation of phytase dosing is not a new strategy and so-called ‘super dosing’ approaches have been used since around 2010. However, in many cases an elevation in phytase dosing has been explicitly geared toward FCR reduction and often the increase in phytase concentration is done without any formulation changes. A recent feed formulation analysis by dsm-firmenich using current feed ingredient prices (as of April/May 2022) and a typical European broiler grower diet, shows that increasing the dose of RONOZYME® HiPhos from 500 to 2000 FYT/kg can generate an additional EUR5/metric ton in feed cost saving. This feed cost reduction is achieved largely by displacement of an additional 3kg/metric ton of inorganic phosphate. Articulation of such strategies with the use of NIR technology to monitor dietary phytate concentration is recommended to ensure adequate substrate supply to carry a more aggressive phosphorus matrix.
2. Enzyme combinations have significant potential to reduce feed cost and matrices for enzyme admixtures can be applied safely when following guidance on sub-additivity across enzyme classes. A recent feed formulation exercise by dsm-firmenich, again using a standard European broiler grower diet and updated feed ingredient pricing, revealed that an additional feed cost reduction of EUR11/metric ton could be achieved (in addition to the value described above). Specifically, the use of a combination of RONOZYME® WX (energy matrix only), RONOZYME® ProAct (digestible amino acid matrix only) and RONOZYME® HiPhos (adding a conservative amino acid and energy matrix on top of the approach detailed above) reduced feed cost from EUR543/metric ton to EUR532/metric ton. These feed cost savings were reinforced by a similar feed formulation example using a standard Asia-Pacific broiler grower diet. Here, the use of a combination of RONOZYME® ProAct, RONOZYME® HiPhos (2000 FYT/kg) and RONOZYME® Multigrain generated a USD32/metric ton of feed cost reduction compared with a formulation based on 1000 FYT/kg RONOZYME® HiPhos with only a mineral matrix applied. These reductions were mostly associated with a reduction in dietary crude protein and added fat sources and an increase in the inclusion of lower cost feed ingredients such as wheat bean, oat hulls and other locally sourced raw materials. The value of energy matrices here is also substantial as the cost of 10 kcal/kg of metabolizable energy today is EUR1.5-4.0 depending on the diet, animal species and geography.
Conclusion
Feed enzymes are an old friend to commercial nutritionists. Use of these technologies reduce nutritional input costs, bring more flexibility in least cost formulation, support animals’ health status and improve and sustainability of production. During these times of unpredictable volatility and catastrophic raw material prices, the value of feed enzymes as part of the nutritionist’s toolbox has never been higher. The effects are consistent, measurable, scientifically credible, and economically valuable. As we navigate the uncertain future together, please consider refreshing your approach to feed enzyme application to further optimize the value that this remarkable technology can bring.
Please speak with your local dsm-firmenich representative for more information and advice.
18 May 2022

